�����ӳ�����ԭ�������ţ�Protozoa�����������ţ�Apicomplexa�������ӳ�٣�Sporozoasida��������Ǹ٣�Coccidiasina���������Ŀ��Eucoccidiorida������������Ŀ��Eimeriorina���������ӳ�ƣ�Cryptosporidiidae���������ӳ�����Cryptosporidium������һ����Ҫ�ij���������ԭ�档�����й㷺�������ף��ɸ�Ⱦ�ˡ������¶��Ұ��������ࡢ�����ࡢ�����240���ֶ��������Ҫ�������������ӳ没�����������ľ�����������������塣�����ӳ没��Cryptosporidiosis����Ҫͨ����-��;����ˮԴ��������ʳ��ȴ�������θ���Ĥ��Ƥϸ�������Ը�к�Ϳ����Ժ�������֢Ϊ�����ٴ�֢״��������䶯�����������½�������������Ӥ��������ȱ�ݻ��߿������ԡ������Լ���(Leitch & He, 2012)��Ŀǰ�ò��ѱ���Ϊ���������6�ָ�к����֮һ���������ӳ��Ⱦ����Ķ�ͯ��к�Ѿ���Ϊ��������״������Ⱦ�ĵڶ���к����Ҳ������1-2��Ӥ����Ⱦ��������Ҫԭ��(Kotloff et al., 2013)��ͬʱ�������ӳ��Ⱦ���̲����ߵ������ʸߴ�61�����ң�Ҳ���շ����̲�������������Ҫ����֮һ(Fayer, 2004)��
�����ӳ�����϶࣬1907���ɽݿ�����ѧ��Ernest Edward Tyzzer�״δ�ʵ��С����������Ƥϸ�����ֳ��壬����1910���������Ϊ�������ӳ棨Cryptosporidium. muris��(Tyzzer, 1910)�������1976�꣬Nime(Nime, et al., 1976)�������о��Ŷ��ױ������˵������ӳ没��ֱ��1982�������ѳǺ���ɼ���Ĵ�����Ⱦ�������֣��ұ��ֳ��밬�̲�������شΣ��ò���ʼ����ҽѧ��Ĺ㷺��ע(1982)��1993��(Mac et al., 1994)������˹�������ܶ��ֻ��г�����ʷ�������������ص�һ���ɳ�������ˮˮԴ��Ⱦ���µ������ӳ没�����������ӳ�Ĺ�����������������һ���µĸ߶ȡ��й������������ӳ没����1987�꺫����(�����ȣ� 1987)���Ͼ��з��֣�������ҹ�ɽ�������ϡ����������ɡ����ա����ϵ�ʡ��½���������������ŷ������в�ѧ�о������IJ������ƣ�Ŀǰ�����ˡ����鶯����ࡢ���������������ڼ�����27����Ч�֣�40���������(Fayer & Santin, 2009; Fayer et al., 2010; Plutzer & Karanis, 2009; Ryan et al., 2014)�����������������ӳ���6�����ϵ���Ч�֣��ֱ�ΪС�����ӳ棨C. parvum�����������ӳ棨C. meleagridis�����������ӳ棨C. muris�����������ӳ棨C. suis����Ȯ�����ӳ棨C. canis����è�����ӳ棨C. felis���������ӳ���һ��ˮԴ�Բ�ԭ�������������ų��������ˮ���д�����£�ˮԴ�Դ���ĿǰҲ�������ӳ没��������Ҫ;��֮һ��������Ṥҵ�����̵��ƽ������ݼ�����������ĽӴ��������У����ɸ�Ⱦ���ų��������ӳ�������Ⱦ����ˮ�����ֳ���ˮԴ��������������Ⱦ�����۶�������ȱ�ݸ��廹������������˵����һ�����ݺ��ӵĹ�����������(Baldursson & Karanis, 2011; Chalmers, 2012)�����ľͽ������������������������ӳ�IJ�ԭѧ���������������в�ѧ���²��ԡ����ͷ��εȷ�����о���չ��һ������
1 ��ԭ
1.1 ����ѧ����
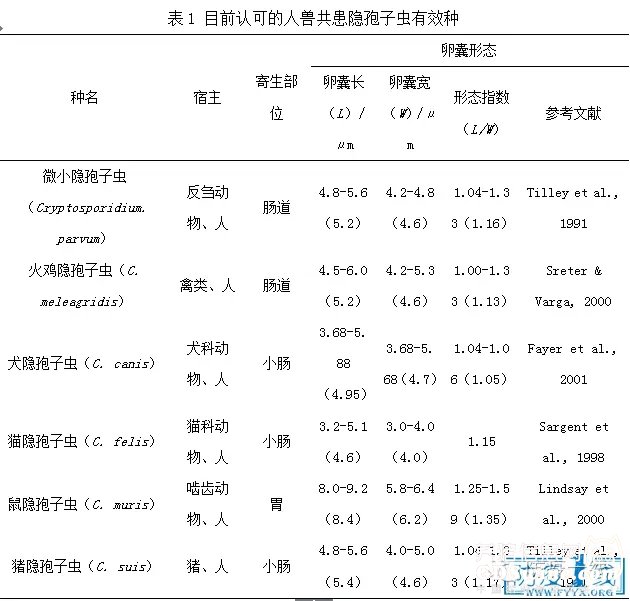
Ŀǰ���Ͽɵ�6�������������ӳ���Ч�ְ���С�����ӳ棨C. parvum�����������ӳ棨C. meleagridis�����������ӳ棨C. muris�����������ӳ棨C. suis����Ȯ�����ӳ棨C. canis����è�����ӳ棨C. felis�������������ӳ�������̬�������ƣ���ΪԲ�λ���Բ�Σ��ұڹ⻬���������ѷ졣�����ų�������Ϊ������ң����������ұڣ��ڲ���Լ�ɼ�������¶���㽶�������ӣ����������ڿ��Կ������塣��������Ⱦɫ�����ӳ����ҳ�õ���ɫ�����������λ������Σ��ɼ��ϱ���Ĥ�������εĿ��ݡ���ͬ�ֵ������ӳ��С��������λ����һ���IJ�𣨼���1����
��Ȼ��ĿǰΪֹ��������ԭ�������ӳ���Ȼ�������������Ŀ�����ǻ���������������о�������ʾ�������ӳ�ͬ������������Ա�繭�γ桢�����ӳ�������ӳ���ںܴ�IJ����ȱʧ�˶�����ϸ�����Լ�����������������(Abrahamsen et al., 2004; Xu et al., 2004; Zhu et al., 2000)������֮�⣬�������кܶ���ص�����������������������������ϸ����������У����ɳ����Ĥ�����ϣ��γɶ��ص�Ӫ���������٣����ڱ������Һͺ������������̬��ǰ�߿���ʵ�ָ�Ⱦ�����ڵ��Է���Ⱦ���������ʷ��������̬��С��ͬʱȱ�������ҡ���������Ĥ����̬�ṹ�������е����п����ҩ��������С��µ��о�����(Koh, et al., 2014; Ryan et al., 2014)���������棬�����ӳ�����ѧ������ԭʼ�Ķ����Ŵس���ӽӽ����������ӳ�����ʷ������ĸϸ�������ⷢ������س�dz�����֮����ͬʱ�������ӳ���¡������س��н��淴Ӧ��Templeton��(Templeton et al., 2010)�������ӳ���س棨Ascogregarina taiwanensis����ȫ���������ݱȽ��о���һ��֤ʵ����ȷʵ���ڽϽ�����ϵ������ϵ��Ȼ���������ӳ桢�س���Ȼ�����������ͬ�Ĵ��������������������ҽΡ�I��֬�����л;������ͪ�ϳ�ø�Լ�һϵ�б��صİ��⵰�ṹ��ȡ����������ӳ�ʹس�����������ϵ������ϵ�������һ�����о������ͣ����������ӳ�����Ĺ�ϵ�������ӳ没�����в�ѧ�����������Ҫ���塣
1.2 ����������
C. parvum, C. hominis ��C. muri���������ӳ��ȫ����������������Ϊ���õ����������ӳ�ķ�������ѧ�����춨����Ҫ���������У�C. parvum��C. muris������������������������塣Windemer��(Widmer & Sullivan, 2012)�о�����C. parvum��C. hominis������DNA���д���95%-97%�����������ԣ�GC����ԼΪ30%��û�д�IJ���/ȱʧƬ�λ����Ե����顣���ߵ�ȫ�����鳤�Ⱦ�Ϊ9.2Mb��������4000������
ĿǰC. parvum��������Ψһһ�����ȫ����װ�������ӳ�����飬��Ӧ8��Ⱦɫ�壬�д�Լ75.3%�ı������ע�͡������ӳ�ͬ�س����ƣ�ȱ�ٶ����壬ȱ������������飬�к˻��������������幦�ܵĵ��ף��������������ѭ��������������֬�����л��������л���̡���л;������ػ����ȱʧҲ��һ���̶���˵���������ӳ��Ӫ����Դ�ܴ�̶�����ֱ�Ӵ�����ϸ���л�ȡ�����Ǵ�ͷ�ϳ����������ǽͽ⡢�������ữ��������л��Widmer��(Widmer et al., 2012)������ֻ��Ⱦ�˵�C. parvum IIc���������C. parvum IOWA�������ȫ������Ƚϣ����ֶ��ߴ���һ���ָ߶ȷ���Ļ�����C. parvum IIc���C. hominis����������������C. parvum IOWA�ꡣ�Ʋ�����C. parvum IIc���C. hominis���߶�ֻ��Ⱦ�ˣ������ڳ��ڵĽ����������γ����ԶԲ�ͬ������������Ӧ��Ϊǰ��Ľ���ģʽ��
C. murisȫ�������ͼ��2008�깫��������C. parvum��C. hominis �ڻ������С������������Լ��������ϻ������ƣ������������ؼ����죺������������һ���ɺ˻�������������ѭ��ø��ATP�ϳ�ø������������ữ����(Mogi & Kita, 2010; Widmer & Sullivan, 2012)��C. muris��Ȼ��C. parvum��C. hominisһ��ȱ������������飬��������س�һ������������Ľṹ�͵��ס�����һ���̶���֤���������ӳ�ʹس���ܴ��ڹ�ͬ�����ȣ��������ڶ�����������ṹ�͵��ף�C. parvum��C. hominis�������ȱʧ�Ƿ����ڴ�C. muris �ֻ������Ժ������ɵ�(Widmer & Sullivan, 2012)��
���������ӳ�Ĺ��ܻ�����ѧ�о���ԱȽ��ͺ����������ӳ渴�ӵķ�����ȱ���ȶ���תȾϵͳ�Լ���Ч����Դ����������ء����ŷ�������ѧ�����Ľ�������Щ���⽫�����ơ�Li��(Li et al., 2014)����������ø�鵼�ĵ��ؼ����ɹ�����ɫӫ�⵰�ͺ�ɫӫ�⵰��תȾ�������ӳ����ڡ�Beverley ��(Beverley, 2015)����Crispr cas9�����ɹ�ʵ���˶������ӳ������ı༭��Ϊ�µĹ��ܵ����о��ṩ���ȶ��ļ���֧�ţ�ʵ���������ӳ湦�ܻ�����ѧ�о����·�Ծ��
1.3 ϵͳ���༰����������
���������ӳ�ķ�������������ȱ��ԭ������һ���Թ�������ϵ¶��Ŵ��ּ䷱��������ԭ�������Բ���������ѧ������ڷ�������һֱ���ںܴ�����顣���ŷ�������ѧ�о������ķ�չ����Ҫ���ݳ���������̬�������λ�����������仯���ٴ�֢״�ȵĴ�ͳ����ѧ�����Ѳ��������������ӳ���ּ���ࡣĿǰ���ձ��Ͽ�Xiao��(Xiao et al., 2004)��������������ӳ����ֵĻ���������������������Ϣ���������ҵ���̬ѧ��ֵ���ϡ��Ŵ�ѧ�������ṩ�����λ����Ŵ���Ϣ��������֤����Ȼ��Ⱦ�����������Ⱦ�����������ԡ����ع��ʶ���ѧ��������ICZN���������������ӳ��ǹ��ɹ�������������в����Ҫ���أ�Ŀǰ���Ͽɵ�6����Ч�������Ƕ�����ȱ�ݸ��廹���������������ɸ�Ⱦ������C. parvum �����Ҳ�DZ����������ࡣ������������ӳ���ֳ�һ�������������ԣ������Ƿdz��ϸ�����C. parvum��������II�ͱ��ֳ��dz��������������ԣ����Ը�Ⱦ��ţ���ˡ������ڶಸ�鶯��ʵ��������������Ⱦ���෴��C. meleagridis, C. canis, C. felis��C. muris ���ֳ���Խ�ǿ�����������ԣ����˸�Ⱦ���⣬�ֱ��Ⱦ�ݡ�Ȯ��è����(Fayer, 2004)�������о���Ϊ��C. hominis ���зdz�ǿ������������ֻ��Ⱦ�ˣ���Morgan��(Morgan et al., 2000)���о����֣�C. hominis ��ɸ�Ⱦ�����鶯�����ޣ��Ӷ�����������רһ�ԡ����⣬�о�������ʵ����������C. hominis���ɸ�Ⱦ��������Сţ�����Dz���Ⱦ��������С��Xiao��Ryan��(Ryan, 2014; Xiao, 2010)�о�����C. parvum����������IIc����ֻ�ܸ�Ⱦ�˵ģ����ֳ���ǿ�����������ԡ�
2 ����ʷ����������
2.1 ����ʷ����
��������������ӳ涼�ǵ�һ�Ե�ϸ���ڣ������⣩�����棬���з����ξ���һ��������Ƥϸ��ˢ״Ե�Ĵ�������ڽ��У�������ֳ��ֳ��������ֳ��������ֳ�����Σ�ͼ1��������治ͬ���ǣ������ӳ��������ֳ�������ڽ��У���������ӻ������к�ڡ������������ͣ���������ֱ�����������ҽ�����һ�ַ�����ʵ�ֳ��������Ը�Ⱦ����������ų������Ⱦ����������Ϊ��Ⱦ�Ρ����ӻ�������Ψһ�������ԽΣ������ų����⣬�����˵���������֮�����������Ҳ������ˡ���θ����������ĺ�������Ƥϸ�������ұڷdz���̣����Ա����ڲ��������ӵ�����������Ӱ�졣Dumeter��(Dumetre et al., 2013)ͨ��ԭ���������о��������ұ����������ͨ�����ϲ��ʡ����ҽ������ں�����������ҹ��̣����Ұ���һϵ�����ص��������绹ԭ������������̼���¶ȡ���ø�����εȡ����ұ�һ�˵ķ�϶�ѿ���������������������������ݳ�������������������Ƥϸ�����ڱ�����İ�Ĥ������ʼ��γ��ɳ���ݣ������ڿ����ڿ�ʼ������ֳ���ȷ���Ϊ�����壬��3�κ˷��ѷ���Ϊ��һ����ֳ�壬����ĵ�һ����ֳ�庬��8����ֳ�ӣ���ֳ�ӱ��ͳ�������������Ƥϸ��������Ϊ�ڶ�����ֳ�塣�ڶ�����ֳ�徭�˷����γɺ���4����ֳ�ӵĵڶ�����ֳ�壬�в���һ�ϴ�IJ��塣��ֳ�ӽ�һ������Ϊ��С�����塣С���������4�η��ѣ��γ��ӵ�ͷ״��16��С���ӣ��ޱ�ë�Ͷ��壬������ǰ����һ���������ܶȺܸߵĸ��������Ʋ����ܾ�����������ء���С���ӽ���ܾ����γɺ��ӣ���������γ��ұں���Ϊ���ң����������ԭ���ʳ���״���̶������ɺ���4�������Ӻ�1������ij�������(Hijjawi, 2010)��
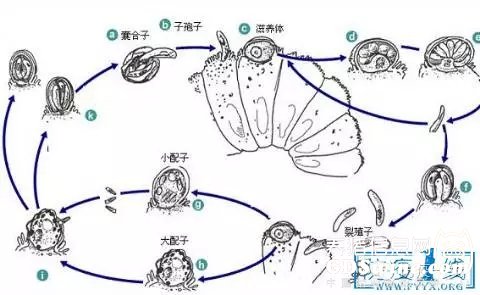
ͼ1 �����ӳ�����ʷͼ��
�����ӳ���������ʷ�ڴ������������3-5d������ɣ��������߹��������ĸ��壬��Ⱦ��������ʱ�����1-���ܣ�����ֻ�Ǽ�Ъ�Եļ����ң���������ȱ�ݸ��壬�����ֳ����ԡ����ڵĸ�Ⱦ���ɳ�������֮�á������ӳ�İ�����Ⱦ������ID50���ܵͣ�ͨ������Ը����������ͬ������C. parvum���ҵ���Ը�߸�Ⱦ�о����֣�UCP���ID50��9-1024������֮�䣬��IOWA���ID50Ϊ87������(Teunis et al., 2002)������ǰ�ķ�ۻ������ֳ��ϸߵ����ԡ���Ⱦ�����ų�����������Ҳû�й̶�����ֵ���˹���ȾСţ105�����ң�7-10d���ų�109-1010����
����2002��Hijjawi��(Hijjawi, 2002)�ͷ����������ӳ�����ʷ�д���һ������֮��������س����Ƶ�����ĸϸ������Gamont-like��ϸ���ⷢ���Σ���Щ���ڶ��о�ѧ��(Hijjawi et al., 2004; Huang et al., 2014; Koh, 2013; Koh, 2014)��ʵ�����ݲ���֤������һ����ȷʵ���ڣ������˹���Ⱦ72h���С����ģ���д�����������ĸϸ������������ε���Դ�����������в���ȷ���Ʋ���Ϊ������δ�ɹ���������ϸ������������ϸ���ⷢ���������(Rosales, 2005)��
2.2 ��������
�����ӳ����������һֱ����������Լ�����ӳ�����ѧ����ԭѧ������ѧ�Լ�ҩ��ɸѡ�о���չ����Ҫ���ء������������������³����ԵĻ�����ң�����ʱ��ı������ң�����ͨ����ʵ�鶯�ﶨ�ڶ��䴫�����֣����ںܴ�̶�����Լ�������ӳ��о��ķ�չ��6����Ҫ�����������ӳ����о���Ϊ�������Ȼ��C. parvum��Ӱ�����������������ذ������ҷ�����ʱ�䡢����ϸ�����͡����������ȡ�����������������ڼ����Ͻ��еģ�����C. parvum��C. baileyi�����������ӳ棩���ɳɹ��������������ʷ���������ڴ���֯�к��ѽ������ӳ�ĸ��������ηֿ�����һ���̶�����������������ģ�͵�ʹ�÷�Χ(Current & Long, 1983; Hijjawi, 2010) ���̶��ڶ��о�ѧ�߲���̽��ϸ������ģ�ͣ�Ŀǰ�õ��������˵Ľ�ä��ϸ����HCT-8��������ϵ, ���������ϸ��ϵ�����ϸ��ϵ�����������ȫ������ʷ������ң������ҵIJ������dz��ͣ��Ҳ��ܳ������������������Щ����о���֣�C. parvum ��HCT-8ϸ�������������Խϴ���ϸ���ͣ��ҳɹ����ֵ���������ں���û�з������Ʋ�Ҳ������ϸ�����������йء�Hijjawi��(Hijjawi,2010)��Karanis(Karanis & Aldeyarbi, 2011)����������ͨ������������ϵPHֵ��٤�����߷��յȷ����������ӳ������ʱ���ӳ���25d��King��(King, 2011)ͨ�����������������������������Һ�������ӽ������ĸ������ٸ�Ⱦϸ�����ɽ���Ⱦ�����4����Castellanos-Gonzalez��(Castellanos-Gonzalez, 2013)���ô��˳���Ƭ���з��뵽��������������Ϊ�������壬ͬʱ�����������ӡ� ���������ӹ��������ɽ������ӳ�����������ӳ���120h����Щ�Ľ����������ӳ���ϸ��ƽ̨�Ͻ���ҩ��ɸѡ�о��춨�˻�����
����������ӳ����������ε���ϸ������ϵͳ��Hijjawi��(Hijjawi et al., 2004)��2004���״�����ġ�������һ������ϵ�����ڸ�Ⱦ��8d��������ɵ����ҡ���ȻGirouard��(Girouard et al., 2006)ʹ�����Ƶ���Ѫ��������ϵȴû���ظ�����һ�о�������������������о�ѧ��(Hijjawi et al., 2010; Koh et al., 2013; Zhang, 2009)֤ʵ������ϸ��������ϵ�У������ӳ�Ļ�����DNA���������ӵģ����Ҳ�ͬ�ķ������磺�����ӡ������塢��ֳ�塢С�����塢���������ĸϸ����ϸ����Ҳ����ͨ��ɨ��羵������ѧ����������Ĥ�м�����������һ���̶���֤ʵ�������ӳ���ϸ������ϵͳ�Ŀ����ԡ�Yang��(Yang et al., 2015)�Ƚ��о���ϸ��������ϵ����ϸ��������ϵ�������ӳ治ͬ�����ε�����ѧ�������εĻ�����������죬��������������ϵ�������ӳ�Ļ�������������ƣ�������ϸ��������ϵ�У�һЩ����ı�������ӳ�ЧӦ�ұ�������Ե͡�����������������ϵ�������ӳ�������������ں��ֲ��컹��Ҫ��һ���Ĵ����о���֤ʵ��������ϸ��������ϵ�Ľ����������ӳ淢������ѧ��ҩ��ɸѡ�о�ȷʵ������̱�ʽ�����壬����ȫ�����˶���ģ�ͻ�ϸ������������
���ߣ���꣬�����ɣ��Ž��W*������ޣ�����Ȩ�������ȣ�����ɽ�������
���ߵ�λ���㶫ʡũҵ��ѧԺ���������о������㶫ʡ�����߲������о��ص�ʵ���ң��㶫ʡ��ҽ������������ʵ����
��ѡ�ԡ�����һ�ߡ�2016���4����47��
| �鿴�������� �������� |
|
|
| �������� | ||||
|







